研究内容(小口)
植物をとりまく環境は空間的にも時間的にも多様である。これに対し動くことのできない植物は、それぞれの環境において効率よく光合成・成長を行うために様々な性質を変化させている(順化・馴化)。しかし、植物によってその順化能力は大きく異なることが知られており、環境変化後の適応度に強く影響していると考えられている。どのような植物がどこまで順化できるのだろうか?この問いは、将来の環境変動に対する生態系の応答を考える上でも重要であり、本グループのメインテーマである。
詳しくはグループのWebpageにて(リンクはこちら)。
① 葉の解剖学的性質と順化能力の関係
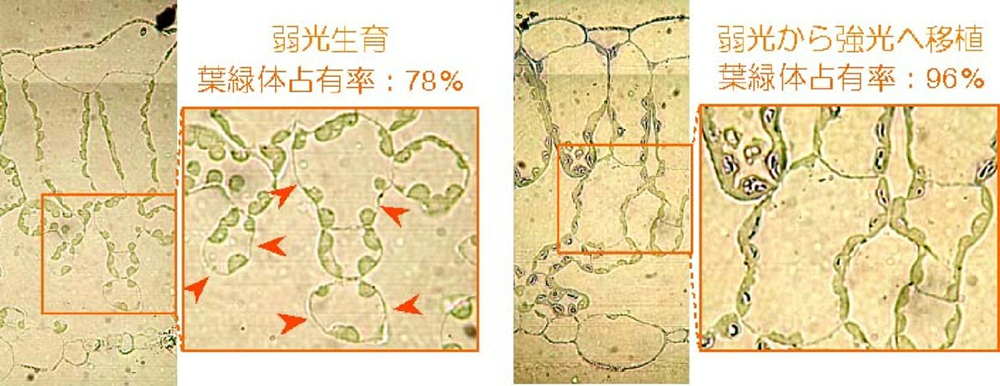
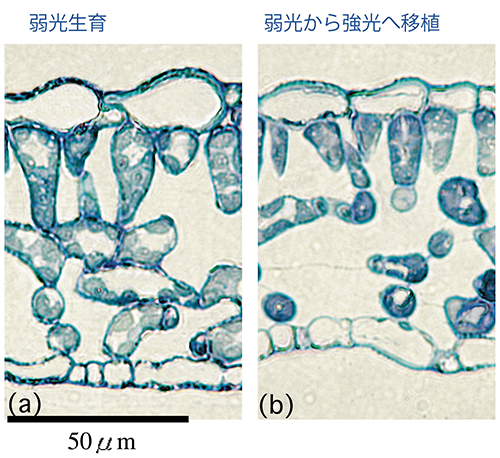
葉緑体は、光合成の基質であるCO2の拡散抵抗を減らすために、葉肉細胞表面付近に位置する必要がある。細胞間隙のような気相中に比べ、細胞内のような液相中の拡散抵抗は10000倍高いためである。もし葉緑体が細胞表面から離れれば、CO2の供給不足がおき、光合成の効率は著しく低下する。そのため、高い光合成能力を持つために葉緑体を増やすには、葉肉細胞が細胞間隙に接する面積を増やす必要があると考えられており、この形態的な制約が植物の順化能力に強く影響していると考え、研究を行なっている。例えば、弱光生育のシロザの葉では葉肉細胞表面を葉緑体が完全に占有してはいなかったため(78%)、強光へ移したあとにその部分を葉緑体が占有するように大きくなることで光合成能力の上昇が起きたこと(図1)、一方でブナでは、弱光生育の葉の葉肉細胞表面にほとんど隙間が無く、葉の厚さを変えることもなかったために、光合成能力が増加しなかったことを明らかにし(図2)、葉の解剖学的性質が順化能力を制限することを示した。林冠木を倒して林床の光環境を変える野外操作実験や、種内変異を利用した順化能力に関わる遺伝的要因の解明などを進めている。
② 光阻害のメカニズム・光阻害耐性の決定要因
光は植物にとって欠くことのできない資源である一方、そのエネルギー故に光合成器官に損傷を与える。この光阻害と呼ばれる現象は、光合成速度の低下を介して植物の成長速度の低下につながる。暗い環境で生育していた植物が明るい環境にさらされるようになった時に、上記のシロザのように光合成能力を高められる種がいる一方で、光合成能力を低下させてしまう種が存在するのは、光阻害が原因の一つと考えられる。ほかにも、低温や乾燥などのストレスが高まると、光阻害が強まることも知られており、植物の環境応答に関わる重要な要因の一つである。光阻害の初期には光化学系IIの活性が低下することが知られているが、そのメカニズムはまだ熱い論争が続いている。また最近は変動光下で光化学系Iも光阻害を受けることが報告され、研究が活発化している分野である。本グループでは、異なる種やエコタイプを用いて、阻害光の光強度・波長を変えたり(図3)、葉の内部の光強度の勾配に注目して、光阻害実験を行い、そのメカニズムおよび光阻害耐性の決定要因を調べている。

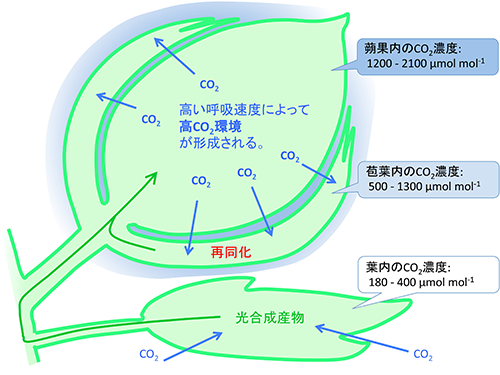
③ 将来の高CO2・高温環境に対する植物の応答
大気中のCO2濃度は上昇を続けており、100年後には現在の2倍の濃度になる事が予想されている。CO2は光合成の基質であるため、植物の成長は促進される事が期待されるが、その応答は種によって異なり、しばしば光合成能力が低下する事で成長促進が抑えられることが報告されている。そのため、どのような形質をもつ事が高CO2環境での持続的な成長促進につながるのかを調べるため、様々な研究が行われてきた。これまでの高CO2応答に関する研究はほとんどが、Growth chamber, Open top chamber (OTC), Free Air CO2 Enrichment (FACE)といった人工的に作り出した高CO2環境に現在のCO2濃度に生育する植物を移して応答を見るという短期応答を観察するものであった。高CO2環境に置いた植物はその直後は光合成速度が上昇するが、その後、炭水化物の蓄積が起きて光合成能力が低下してしまう例が多く報告されているように、この短期応答が長期的な高CO2環境での応答であるとは限らない。本グループでは植物個体内で呼吸によって高CO2条件となっている環境(繁殖器官など)という植物が長年保ってきた高CO2微環境への葉緑体や器官の応答(図4)、および、様々な環境に適応したエコタイプの形質とエコタイプ間の高CO2応答の差の関係に着目して研究を行っている。